


Cards In This Set
| Front | Back |
|
How many tissues in the early gastrula have their fate determined?
|
One- the dorsal lip
|
|
Re spemann, what happens to newt embryos when the zygote is constricted in the plane of the first cleavage?
|
At the 16 cell stage a nucleus from one side escapes to the nuclear free half, and then proceeds to begin cleavage divisions on this second side. Eventually, two normal embryos form.
|
|
Re spemanns constriction along the plane of first cleavage: what did this show?
|
That the early amphibian nuclei are the same, and that each is capalbeof forming an entire embryo.
|
|
What happens when the grey crescent is localised in only one of the two blastomeres after the zygote is constricted perpendicular to the plane of first cleavage?
|
Abnormal differentiation of the blastomere which does not recieve the grey crescent. Ectoderm, mesoderm and endoderm are still present but no dorsal structures are, leading spemann to term this mass of cells the "belly piece"
|
|
What structure does the grey crescent lead to and what function does this structure have in gastrulation?
|
Dorsal lip, committed to invaginate the blastule (and thus initiate gastrulation i think)
|
|
What creates the nieuwkoop center?
|
He Nieuwkoop center is created by the cytoplasmic rotation that occurs during fertilization (see Chapter 7).
When this rotation is inhibited by UV light, the resulting embryo will
not form dorsal-anterior structures such as the head or neural tube
|
|
What is the product of the Xbra (brachyury) gene?
|
The Xbra protein is a transcription factor that activates the genes that produce mesoderm-specific proteins.
|
|
Xenopus early fate map
|
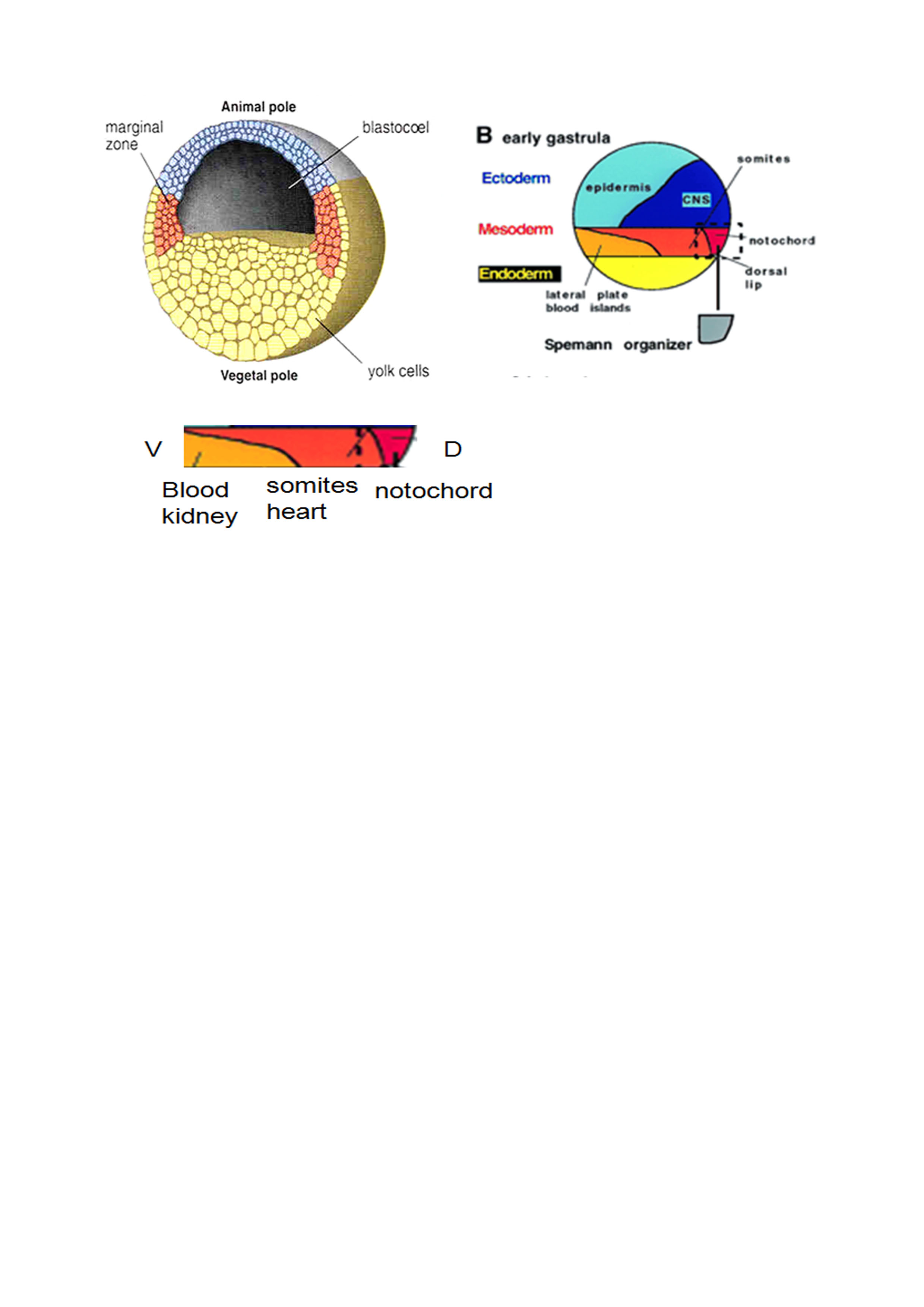 Animal pole – epidermis and central nervous system Vegetal pole – endoderm (gut, lungs) Marginal zone – mesoderm |
|
What induces cells overlying the vegetal cells to begin expressing the transcription factor Xbra (brachyury) which specifies mesodermal change by activating genes that produce mesoderm specific proteins?
|
Vegetal
cells secrete TGFbeta like factors such as Vg1
and VegT, a transcription
factor that instructs the endoderm to secrete activin
and Nodal proteins (all TGF-b family members).
|
|
Spemann organiser overview
|
The organizer is a group of cells that is fated to become
dorsal mesoderm, and that can “instruct’ overlying tissues about how to behave.
The steps in the formation of the organizer are to:-
1) tell the cells that they are dorsal, and
2) to tell the cells that they are mesoderm, and
3) to give the organizer cells inducing properties
|
|
How do the spemann organiser cells know that they are dorsal?
|
The beta catenin signal and the nieuwkoop center signals.
|
|
Where does Beta catenin accumulate during early embryo development in xenopus?
|
β-catenin continues to accumulate preferentially at the dorsal side
throughout early cleavage, and this accumulation is seen in the nuclei
of the dorsal cells ; Schneider et al. 1996; Larabell et al. 1997).
This region of β-catenin accumulation originally appears to cover both
the Nieuwkoop center and organizer regions. During later cleavage, the
cells containing β-catenin may reside specifically in the Nieuwkoop
center (Heasman et al. 1994a; Guger and Gumbiner 1995).
|
|
What is beta catenin required for?
|
β-catenin is necessary for forming the dorsal axis, since experimental
depletion of β-catenin transcripts with antisense oligonucleotides
results in the lack of dorsal structures
|
|
What occurs when exogenous Beta catenin is injected in the ventral side of the developing embryo?
|
The injection of exogenous β-catenin into the ventral side of the embryo produces a secondary axis. β-catenin is part of the Wnt signal transduction pathway and is negatively regulated by the glycogen synthase kinase 3
|
|
What are the molecular mechanisms that lead to the expresion of Beta catenin in the dorsal, but not the ventral, parts of the developing embryo?
|
GSK-3+axin+APC complex binds Beta catenin )(armadillo) and degrades it via phosphorylation. However, dorsally, Dishevelled binds GSK-3, allowing beta catenin to accumulate.
|
